Synergistic Effects of L-Arginine and Metformin on Oxidative Stress, Inflammation and Glucometabolic Enzymes in Diabetic Rats
-
Saka Waidi Adeoye
Department of Physiology, College of Health Sciences, Ladoke Akintola University of Technology, Ogbomoso, Oyo, Nigeria
Igbayilola Yusuff Dimeji

Department of Human Physiology, College of Medicine and Health Sciences, Baze University, Abuja, Nigeria
Lawan, Hamidu JabbaDepartment of Human Physiology, College of Medicine and Health Sciences, Baze University, Abuja, Nigeria
Osukoya EmmanuelDepartment of Physiology, College of Health Sciences, Ladoke Akintola University of Technology, Ogbomoso, Oyo, Nigeria
Abaloja Temiloluwa SamuelDepartment of Physiology, College of Health Sciences, Ladoke Akintola University of Technology, Ogbomoso, Oyo, Nigeria
Ogunsina Damola OluwakemiDepartment of Physiology, College of Health Sciences, Ladoke Akintola University of Technology, Ogbomoso, Oyo, Nigeria
Zakari, MuhammedDepartment of Human Physiology, College of Medicine and Health Sciences, Baze University, Abuja, Nigeria
| Received 16 Aug, 2024 |
Accepted 24 Sep, 2024 |
Published 25 Sep, 2024 |
Background and Objective: Diabetes is a complex metabolic disorder characterized by impaired glucose regulation, leading to glucose intolerance and hyperglycemia. This study explores the effects of L-arginine supplementation on inflammation, glucometabolic enzymes and oxidative stress biomarkers in male Wistar rats with alloxan-induced diabetes. Materials and Methods: The 50 adult male Wistar rats were divided into 5 groups of 10. Group A (control) received normal saline. Group B was induced with diabetes using alloxan. Groups C, D and E received alloxan plus metformin, L-arginine, or both for six weeks. Inflammation, oxidative stress and key glucose-regulating enzymes were assessed post-study. The data were presented as Mean±Standard error. One-way Analysis of Variance (ANOVA) was employed to assess statistical differences between groups, followed by Tukey’s post hoc test for individual comparisons. A 95% confidence interval was applied and results with p<0.05 were deemed statistically significant. Results: The combination of L-arginine and metformin significantly lowered blood glucose levels compared to the diabetic group. The combination therapy also decreased malondialdehyde levels, increased superoxide dismutase and catalase activity and reduced inflammatory markers like TNF-α, IL-6 and C-reactive protein, indicating improved glucose metabolism over individual therapies. Conclusion: In male Wistar rats with alloxan-induced diabetes, the combined administration of L-arginine and metformin significantly reduces inflammation, oxidative stress and dysregulation of glucometabolic enzymes. These findings suggest that combination therapy may be an effective strategy for managing diabetes and its complications.
| Copyright © 2024 Adeoye et al. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
Diabetes is a severe health condition around the world. It produces excessive blood sugar levels, which can lead to sickness and even death. In 2019, around 463 million people globally have diabetes and this figure is predicted to rise to 700 million1. Diabetes is multifaceted, with a complicated interplay betweengenetic, environmental and lifestyle variables. Among the various forms of diabetes, alloxan-induced diabetes in rats has been widely used as a model to study pathophysiology and possible interventions because it is reproducible and resembles certain aspects of human diabetes2. Innovative medicines to control the consequences of diabetes are desperately needed, given the disease’s widespread prevalence. Because of its vasodilatory properties and ability to enhance vascular function in diabetes, L-arginine, an amino acid precursor to nitric oxide, shows promise as a therapeutic intervention3. According to recent research, administering L-arginine over an extended period of time may improve insulin sensitivity in individuals with type 2 diabetes, indicating its potential as a novel therapeutic approach4. The L-arginine may provide a major means of controlling type 2 diabetes problems globally by addressing vascular dysfunction.
In recent years, researchers have explored the therapeutic potential of dietary supplements and L-arginine has emerged as a promising candidate. The L-arginine, a semi-essential amino acid, plays a crucial role in many physiological processes, including the synthesis of nitric oxide (NO), a molecule with vasodilator and anti-inflammatory effects5. Nitric oxide is involved in regulating insulin sensitivity and glucose metabolism, making L-arginine an interesting candidate for improving the dysregulation seen in diabetes5.
It is well recognized that nitric oxide (NO) is a multifunctional signaling molecule that helps to maintain cardiovascular and metabolic homeostasis6. Most people concur that diabetes mellitus and chronic hyperglycemia affect the synthesis and bioavailability of NO. Nonetheless, while some research7-11 has shown elevated NO levels in type 2 diabetes people and animal models of diabetes, other research by Habib et al.12, Bahadoran et al.13 and Paulo et al.14 has shown the opposite. In animal models of obesity, hypercholesterolemia and hyperglycemia, NO bioavailability has also been reported to be decreased12,15. Previous studies have investigated the effects of L-arginine supplementation in various experimental models of diabetes. Suessenbacher et al.16 showed that administration of L-arginine improved endothelial function and reduced markers of oxidative stress in type 2 diabetes rats. Similarly, Forzano et al.17 reported improvements in insulin resistance and inflammation in a mouse model of type 2 diabetes after L-arginine supplementation. These findings highlight the potential benefit of L-arginine in mitigating diabetes-related complications. Previously, dietary L-arginine supplementation has been reported to reduce fat mass in Zucker type 2 diabetes fatty rats18, regulated hyperglycemia and dyslipidemia19 and L-arginine and nitric oxide have been shown to have the potential to prevent alloxan-induced beta-cell damage and the development of diabetes and restore the antioxidant status to near normal levels20.
Metformin, a biguanide derivative, mitigates hypoglycemia risk and exhibits anti-hyperglycemic effects, reducing incidence by 31%21. It curtails hepatic gluconeogenesis, heightens insulin-mediated peripheral glucose uptake22 and inhibits adipose tissue breakdown, thereby lowering free fatty acid levels23. Metformin also ameliorates blood pressure and lipid profiles in type 2 diabetes patients24,25. Furthermore, it reinforces antioxidant defenses and activates AMP-kinase, regulating glucose and lipid metabolism26. Metformin usage correlates with reduced macrovascular morbidity and mortality in overweight type 2 diabetes patients, independent of glycemic control27. Additionally, it enhances endothelial function28, restoring microvascular responsiveness in type 2 diabetes rats27 and improving flow-mediated dilatation in normo-insulinemic individuals29.
Dysregulation of glucometabolic enzymes, oxidative stress and inflammation are all linked to diabetes. Metformin and L-arginine are known to affect these pathways separately, but it’s unknown how they work together. Examining their simultaneous effects may reveal antagonistic or synergistic relationships, which could lead to improved treatment approaches for diabetes control. The study hypothesis is that, in male Wistar rats with alloxan-induced diabetes, supplementing L-arginine and metformin together will improve the activity of glucometabolic enzymes and reduce oxidative stress and inflammation more than supplementing either drug alone or in combination with it. The main objective of this study is to investigate the effects of L-arginine supplementation on inflammation, selected glucometabolic enzymes and biomarkers of oxidative stress in male Wistar rats with alloxan-induced diabetes.
MATERIALS AND METHODS
Study area: This study was conducted at the Department of Physiology, Faculty of Basic Medical Sciences, Ladoke Akintola University of Technology, Ogbomos, Oyo state, Nigeria between April to July, 2023. The 50 mature male Wistar rats of average weight between 250-300 g were divided into 5 different groups of 10 rats each; control, L-arginine only, alloxan only, alloxan+L-arginine and alloxan+metformin. These rats had not previously received any experimental manipulations and were judged healthy due to the lack of stress or infection indications. Before beginning the tests, the rats were individually weighed following a 2 weeks acclimatization period. They were housed in well-ventilated plastic cages at LAUTECH’s Department of Physiology, Faculty of Basic Medical Sciences, in Ogbomoso, Oyo State. The animals were fed a regular meal and water on a 12 hrs light/dark cycle, with a temperature range of 25±2°C and humidity of 60-65%. The experimental protocols adhered to the guidelines outlined in the National Research Council’s Guide for the Care and Use of Laboratory Animals.
Ethical approval: The ethical approval was sought and given to conduct this study from Ethics and Review Committee of the Faculty of Basic Medical Sciences, Ladoke Akintola University of Technology, Ogbomoso with a reference number FBMS/AECP/092/23.
Induction of diabetes: Rats were induced into a diabetes state by intraperitoneal injection of alloxan 150 mg/kg dissolved in normal saline after an overnight fast30. The method used to induce diabetes involved injecting 150 mg/kg b.wt., of alloxan intraperitoneally. A 13% alloxan solution was made using regular saline. As 72 hrs after the dose of alloxan. An ACCU-CHEK Active device (Roche Diagnostics GmbH, D68298 Mannheim, Germany) was used to measure glycemia. By puncturing the tail veins, blood was extracted and the selection of type 2 diabetes animals with glycemia greater than 200 mg/dL was made possible by determination of blood glucose level.
Grouping:
| • | Group A (Control group): Were given normal saline throughout the study | |
| • | Group B (DM): Were induced with alloxan (150 mg/kg) and after 72 hrs they were rendered type 2 diabetes | |
| • | Group C (DM+Metformin): Were induced with alloxan (150 mg/kg) as in group B as well treated with metformin | |
| • | Group D (DM+L-arginine): Were induced with alloxan (150 mg/kg) as in group B as well treated with L-arginine (100 mg/kg) daily | |
| • | Group E (DM+L-arginine+Metformin): Were given L-arginine (100 mg/kg) |
All animals had access to a standard diet and water and the experimental procedure lasted for 6 weeks.
L-arginine supplementation and chemical
Dosage and administration: A low dose of 100 mg/kg was used to determine the dose administered to each animal31.
A low dose of 100 mg/kg was used to calculate the dosage given to each animal. Based on the formula:
the computation was made using the rats’ average weight of 250 g. The amount of L-arginine provided was calculated using the knowledge that one 500 mg capsule of L-arginine contains 10 rats weighing 2500 g. The proportion for 2500 g×of L-arginine was estimated using 1000 g of 100 mg L-arginine. As a result, a group of ten male Wistar rats were given 250 mL of L-Arginine. Consequently, each rat was given 25 mg of L-arginine.
Collection of blood and tissue samples
Collection of blood samples: Blood samples were collected after an overnight fast (12 hrs)31,32 after the experimental period. The animals were euthanized using ketamine as a tranquilizer. A vertical incision was quickly made in the abdomen and blood was taken through a heart puncture with a 2 mL needle and syringe, then transferred into standard vials33,34.
Collection of tissue samples: The kidney and liver tissues were harvested, cleaned of adherent tissue and weighed with a sensitive balance (Salter, England). The collected tissues were washed in normal saline to remove blood stains and stored in a standard bottle containing formalin and normal bottles containing normal saline, properly labelled when collected from different groups. The samples were kept cold at -4°C before biochemical analysis.
Assay of C-reactive protein: The C-reactive protein was determined using the Enzyme-Linked Immunosorbent Assay (ELISA) test, employing a rat C-reactive protein ELISA kit from Elabscience, Wuhan, China.
Principle: Each serum reference, control and animal specimen had its microplate well and duplicates were made for each experiment. The unused microwell strips were returned to the aluminium bag, sealed and stored at 2-8°C. Next, pipette 0.050 mL (50 μL) of serum reference, control, or specimen into the designated well. After that, add 0.100 mL (100 μL) of T4 Enzyme Reagent to each well and gently swirl the microplate for 20-30 sec to cover. The sample was then incubated for 60 min at room temperature. The microplate’s contents were discarded either by decantation or by blotting it dry with absorbent paper.
After that, 350 μL of wash buffer was added and decanted (tap and blot). The process was performed twice more for a total of three washes. The directions for using an automatic plate washer were followed. A squeeze bottle was used to fill each well by depressing the container. To decrease variance in reaction times between wells, each well received 0.100 mL (100 μL) of working substrate solution in the same sequence as the reagents. Following that, the microplate was incubated for fifteen min at room temperature. Finally, each well received 0.050 mL (50 μL) of stop solution and was carefully agitated for 15-20 sec. To reduce well defects, a reference wavelength of 620-630 nm was used to measure absorbance in each well at 450 nm using a ELISA reader ((Bio-Rad Laboratories, USA).
Assay of IL-6 (Interleukin-6): The IL-6 was determined using the Enzyme-Linked Immunosorbent Assay (ELISA) test, employing a rat C-reactive protein ELISA kit from Elabscience, Wuhan, China.
Principle: Appropriate number of coated strips were put in the holder. As 50 mL of IL-6 standards, control and specimens were pipetted into the appropriate wells. As 100 mL of the conjugate reagent, ready for use, was added to each well for 10 to 30 sec. A cover was placed on the plate and allowed to sit at room temperature (18-26°C) for 60 min. The liquid was extracted from every well. 3,3 ,5,5 -Tetramethylbenzidine (TMB substrate (100 mL) was added to each well and incubated at room temperature for 15 min. As 50 mL of stop solution was added to each well. To combine the solution, the plate was gently shaken. After adding the stopping solution, after waiting for 15 min the absorbance was measured using an ELISA reader (Bio-Rad Laboratories, USA) set to 450 nm.
Assay of TNF-α (Tumour Necrotic Factor alpha): To mix the solution, the plate was gently shaken. After adding the stopping solution and waiting 15 min, the absorbance was measured with an ELISA reader (Bio-Rad Laboratories, USA) at 450 nm.
To mix the solution, the plate was gently shaken. After adding the stopping solution and waiting 15 min, the absorbance was measured with an ELISA reader (Bio-Rad Laboratories, USA) at 450 nm.
Assay of Caspase 3: The Caspase 3 was determined using the Enzyme-Linked Immunosorbent Assay (ELISA) test, employing a rat Caspase 3 ELISA kit from Elabscience, Wuhan, China.
Principle: A caspase-3 test employing a rat ELISA caspase-3 kit consists of a sequence of methods for quantifying the enzyme’s content in tissue or cell samples. Samples are first prepared by homogenizing tissues or lyzing cells, then centrifuged to extract supernatant. The recovered supernatants are then added to ELISA plates that have been pre-coated with an antibody specific to rat caspase-3. Following incubation, a detection antibody is introduced, which binds to the trapped caspase-3 and forms an antibody-antigen complex. A substrate solution is then added, which reacts with the enzyme-linked detecting antibody to cause a colorimetric change. The colour intensity, evaluated with a spectrophotometer (Thermo Fisher Scientific, USA), is directly proportional to the caspase-3 concentration in the sample. After adding the stopping solution and waiting 15 min, the absorbance was measured with an ELISA reader (Bio-Rad Laboratories, USA) set to 450 nm.
Determination of hexokinase: An enzyme called hexokinase is responsible for catalyzing the conversion of glucose to glucose-6-phosphate. The metabolic process known as glycolysis, which turns glucose into energy, begins with this reaction. The assay for hexokinase quantifies the amount of this enzyme present in a specimen. After preparing and incubating at 37°C for 10 min, a reaction mixture including glucose, ATP (Adenosine Triphosphate), MgCl2 and hexokinase was added. Next, Nicotinamide Adenine Dinucleotide Phosphate Plus (NADP+) and glucose-6-phosphate dehydrogenase were added to the reaction mixture. A spectrophotometer (Thermo Fisher Scientific, USA) was used to measure the change in absorbance at 340 nm. The molar extinction coefficient of NADPH (nicotinamide adenine dinucleotide phosphate hydrogen) and the change in absorbance were used to calculate the hexokinase activity.
Determination of amylase: Amylase is an enzyme that catalyzes the hydrolysis of starch into smaller carbohydrates such as maltose and glucose. The amylase assay measures the activity of this enzyme in a sample. A reaction mixture comprising starch, buffer solution and amylase was created and it was incubated at 37°C for 10 min before iodine was added to the mixture to terminate the reaction. This process was done to measure the activity of amylase. A spectrophotometer was used to quantify the change in absorbance at 620 nm and the amylase activity was computed using the absorbance change and a standard curve.
Determination of pyruvate kinase: The enzyme pyruvate kinase catalyzes the transfer of a phosphate group from phosphoenolpyruvate to ADP (Adenosine diphosphate), resulting in the formation of pyruvate and ATP. The assay for pyruvate kinase quantifies the amount of this enzyme present in a sample. A reaction mixture containing phosphoenolpyruvate, ADP, MgCl2, pyruvate kinase and lactate dehydrogenase was prepared and incubated at 37°C for 10 min. Change in absorbance at 340 nm over time using a spectrophotometer (Thermo Fisher Scientific, USA) was measured and the pyruvate kinase activity was calculated using the change in absorbance and the molar extinction coefficient of Nicotinamide Adenine Dinucleotide Hydrogen (NADH).
Oxidative stress studies: Superoxide dismutase (SOD) activity was assayed in line with the protocol described by Vilela et al.35. Reduced glutathione (GSH) activity was assayed as described by Vilela et al.35.
Catalase (CAT) activity was determined by the protocol of Aebi36 and Malondialdehyde (MDA) level was assayed according to the protocol described by Uchiyama and Mihra37.
Histopathological analysis: The renal and cardiac tissues were dried in ethanol, wrapped in paraffin wax and stored in phosphate-buffered saline (0.1 M; pH 7.4). The liver tissues were prepared and stained with hematoxylin and eosin after being thinly sliced (4 mm thick). Using DPX (Dibutylphthate Polystyrene Xylene) mount, these stained sections adhered to slides permanently38. After that, they were examined under a microscope that was equipped with a camera (Motic Images plus version 2.0) and pictures were captured using the microscope (Olympus Corporation, based in Japan).
Statistical analysis: Data were recorded as Mean±Standard error of the mean. Statistical differences between means were determined by One-way Analysis of Variance (ANOVA). Turkey’s post hoc test was used to identify differences between individual means. Confidence intervals were set at 95% and results with a value of p<0.05 were accepted as significant in all cases (GraphPad Prism 5, GraphPad Software, Inc., La Jolla, California, USA).
RESULTS
Effect of L-arginine on Interleukin-6 (IL-6) in alloxan-induced type 2 diabetes rats: The study’s findings showed that the levels of IL-6 in type 2 diabetes rats (group B) were significantly higher (p<0.05) than in the control group (group A) (Fig. 1). Additionally, the levels of IL-6 in groups C and D were significantly lower (p<0.05) than in the type 2 diabetes rats (group B) following the administration of metformin and L-arginine respectively as shown in Fig. 1.
Effect of L-arginine on Tumour Necrosis Factor-alpha (TNF-α) in Type 2 diabetes rats: The research showed that type 2 diabetes rats (group B) had much higher levels of TNF-α than the control group (group A). Figure 2 group D’s TNF-α level dropped a lot (p<0.05) after getting L-arginine when compared to type 2 diabetes rats (group B). Also, after taking metformin, the TNF-α levels of group C decreased significantly compared to group B’s type 2 diabetes rats following administration of metformin as shown in Fig. 2.
Effect of L-arginine on C-reactive Protein (CRP) in type 2 diabetes rats: The research discovered that the CRP level was much higher in the type 2 diabetes rats compared to the control group (Fig. 3). The CRP level in group D significantly reduced (p<0.05) after L-arginine exposure compared to type 2 diabetes rats in group B. Furthermore, after taking metformin, the CRP level of group C increased significantly (p<0.05) compared to the control rats in group A as shown in Fig. 3.
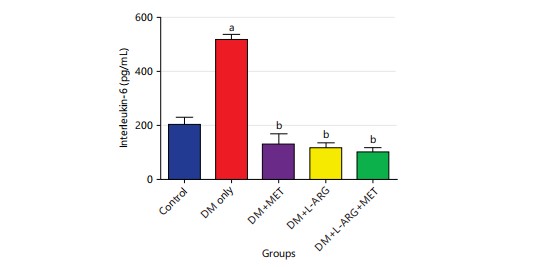
|
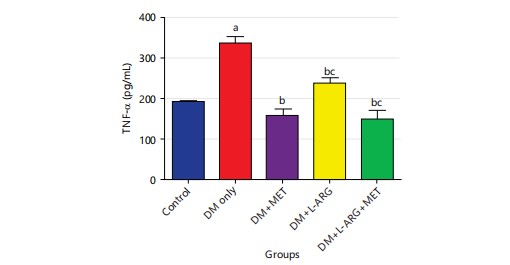
|
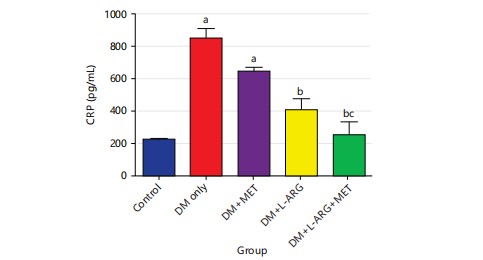
|
Effect of L-arginine on caspase-3 in type 2 diabetes rats: The research discovered that the type 2 diabetes rats had much higher levels (p<0.05) of caspase-3 compared to the control group. Caspase 3’s reduced significantly (p<0.05) after exposure to L-arginine compared to type 2 diabetes rats in group B. Also, the level of caspase-3 in group C increased significantly (p<0.05) after getting metformin compared to the control rats in group A as shown in Fig. 4.
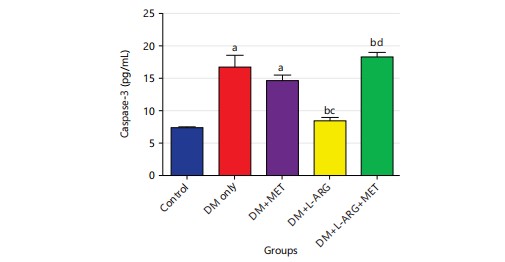
|
|
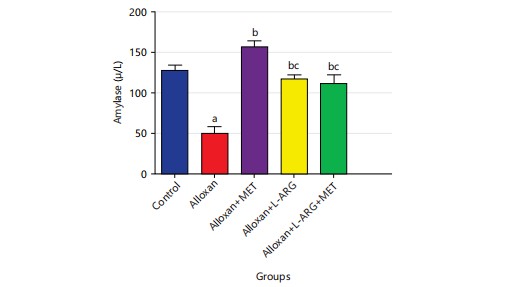
Effect of L-arginine on amylase in type 2 diabetes rats: The research showed that type 2 diabetes rats given alloxan had significantly lower (p<0.05) amylase levels compared to the control group. Furthermore, giving metformin to rats with diabetes (group C) upregulated (p<0.05) amylase levels compared to group B. Also, giving L-arginine to male Wistar rats with diabetes (group D) significantly increased (p<0.05) their amylase levels as shown in (Fig. 5).
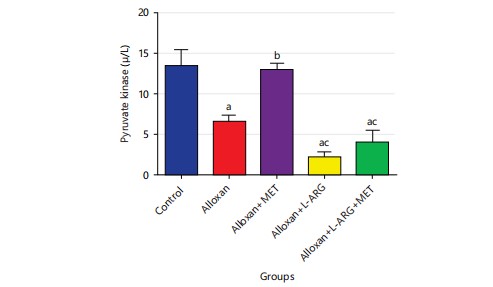
Effect of L-arginine on pyruvate kinase in type 2 diabetes rats: The study in Fig. 6 showed that the pyruvate kinase level decreased significantly (p<0.05) in type 2 diabetes rats compared to the control group. Furthermore, giving metformin to type 2 diabetes rats in group C increased significantly (p<0.05) compared to group B and giving L-arginine to type 2 diabetes male Wistar rats in group D decreased the level of pyruvate kinase significantly (p<0.05).
|
|
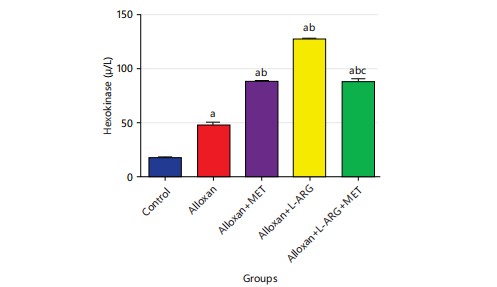
Effect of L-arginine on hexokinase in type 2 diabetes rats: The result showed that the level of hexokinase was significantly (p<0.05) higher in the type 2 diabetes rats that were given metformin, compared to the control group. Furthermore, when metformin was given to rats with diabetes, there was a significant increase (p<0.05) in their blood sugar levels compared to rats given alloxan. Also, giving L-arginine and metformin to male Wistar rats with diabetes caused a significant increase (p<0.05) in their hexokinase levels. These were illustrated in Fig. 7.
|
|
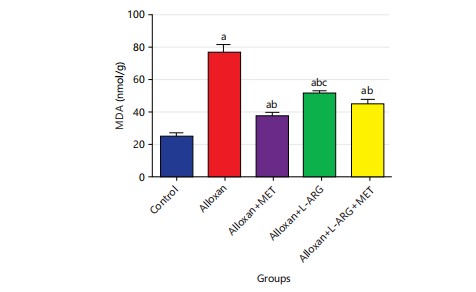
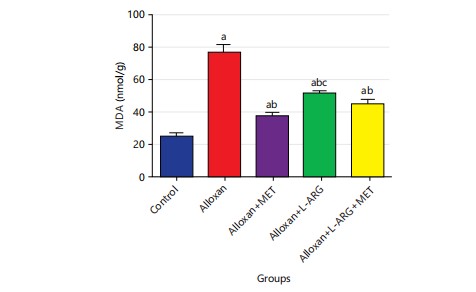
Effect of L-arginine on MDA, CAT, SOD and GSH in alloxan-induced type 2 diabetes rats: The study found that the MDA level significantly increased (p<0.05) in group B compared to the control group. This result showed that the levels of MDA were significantly (p<0.05) lower in the groups that were treated with L-arginine and metformin compared to the group that only received alloxan (Fig. 8).
The study found that the level of CAT decreased significantly (p<0.05) in the alloxan group compared to the control group. Additionally, the study found that there was a significant increase (p<0.05) in the level of catalase in the groups that were treated with L-arginine and metformin compared to the group that only received alloxan (group B). This is shown in Fig. 9.
|
|
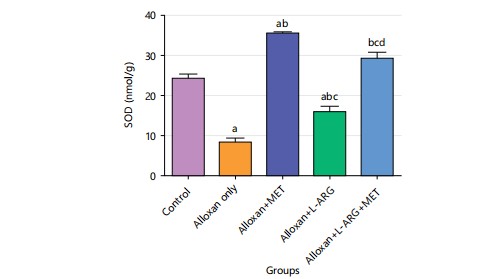
The research showed that in group B, the SOD level was significantly lower (p<0.05) compared to the control group. Also, when comparing the group that only received alloxan (group B), the group that received both L-arginine and metformin showed a much higher level of SOD (p<0.05) as shown in Fig. 10.
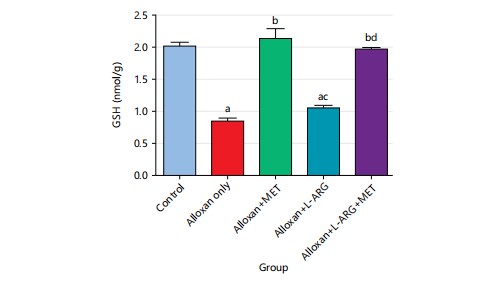
The study found that the GSH level significantly decreased (p<0.05) in the alloxan group compared to the control group. In addition, there was not a significant increase (p>0.05) in the groups given L-arginine, but there was a significant increase (p<0.05) in GSH level in the groups given metformin compared to the group only given alloxan (group B), (Fig. 11).
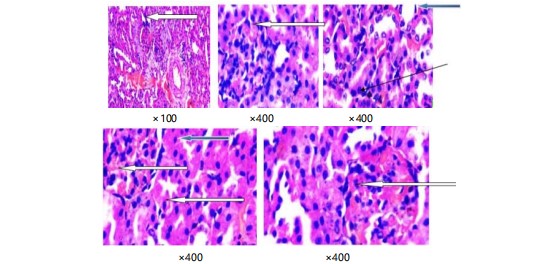
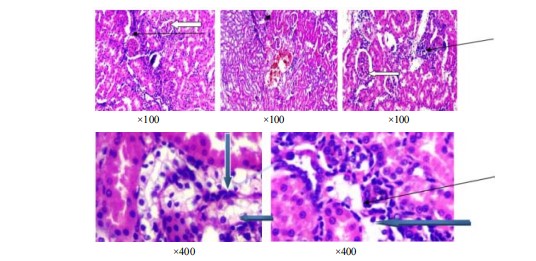
Histological examination of renal tissue in control rats and alloxan-induced diabetic rats: In plate 1, the tubes in the kidney look normal (blue arrow), the outer part of the kidney has normal filter units with normal cells and spaces (white arrow) and the spaces between the kidney’s filters look normal (slender arrow). Pictures of kidney tissues stained with special dyes show the normal structure when seen with a lower magnification of ×100.
Plate 2 shows parts of the kidney that are stained with a special dye. In the pictures, the kidney looks normal in some parts, but in other parts, the kidney cells look damaged and there are a lot of inflammatory cells.
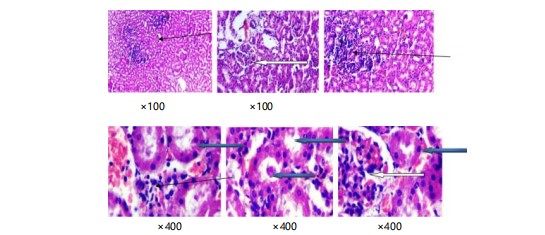
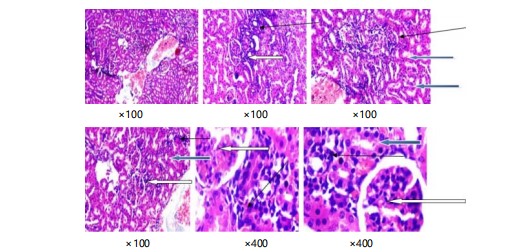
Plate 3 shows the kidney slices stained in purple and red show normal parts of the kidney called glomeruli, with normal cells and spaces around them. The tubes in the kidney also look normal. However, there are some areas where too many inflammatory cells are gathered. Close-up pictures of kidney tissue (plate 4) stained with special dyes that let us see the different parts. The kidney looks normal at a low level of zoom. The outer part of the kidney called the cortex, has normal clusters of cells and empty spaces around them (shown by a white arrow). The tubes inside the kidney also look normal (shown by a blue arrow). The spaces in between the different parts of the kidney look a little bit filled with cells (shown by a slender arrow). There is also a little bit of extra blood in the blood vessels.
|
|
|
|
|
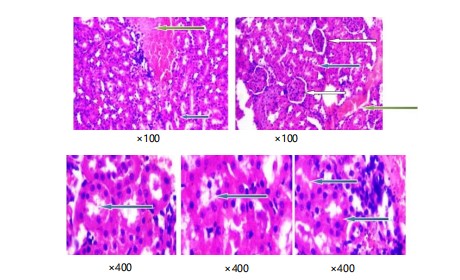
Pictures of kidney tissues stained with Haematoxylin and Eosin (Plate 5) show the normal parts of the kidney. These pictures are taken at a lower magnification of x100. The renal cortex shows normal glomeruli with normal mesangial cells and capsular spaces. The renal tubules also look normal. There is a little bit of congestion in the interstitial space.
DISCUSSION
In type 2 diabetes male Wistar rats, the combined effects of L-arginine and metformin administration on oxidative stress, inflammation and glucometabolic enzyme function were investigated. The results show a number of interesting results. The results of this investigation substantiate the potential anti-inflammatory impacts of L-arginine and metformin supplementation in the context of alloxan-induced diabetes in male Wistar rats. Diabetes is characterized by chronic inflammation, which contributes to insulin resistance and exacerbates the progression of the disease39. Notably, this study identified a noteworthy decrease in pro-inflammatory cytokines, including TNF-α, IL-6, C-reactive protein and Caspase 3, after L-arginine supplementation. These findings were consistent with prior studies6,16. The mechanisms underlying L-arginine’s anti-inflammatory properties are several. The control of NO generation through the activation of endothelial nitric oxide synthase (eNOS) is one well-known method. NO is a powerful vasodilator that has anti-inflammatory characteristics and can decrease inflammatory responses by blocking leukocyte adhesion and migration40. Additionally, L-arginine promotes the production of agmatine and polyamines, both of which are connected to the control of inflammatory signaling pathways41,42. The L-arginine supplementation reduces inflammation via altering polyamine metabolism, which inhibits the activation of Nuclear Factor-Kappa B (NF-κB) and reduces pro-inflammatory gene expression43.
The anti-inflammatory attributes of L-arginine can be ascribed to its role as a precursor for nitric oxide (NO) synthesis. NO, a potent vasodilator, is recognized for regulating immune responses and mitigating inflammatory processes44. The documented decline in pro-inflammatory cytokines implies that L-arginine may regulate the inflammatory cascade in alloxan-induced diabetes, presenting a potential mechanism for its therapeutic effects. Clinical evidence suggests metformin’s potential in modifying inflammation, evident in reduced inflammatory markers like C-reactive protein44-46. Additionally, with combined supplementation, a positive improvement in glucometabolic enzyme activity was observed, suggesting enhanced insulin sensitivity and glucose metabolism. This could be explained by L-arginine’s capacity to increase insulin sensitivity and secretion, as well as metformin’s proven ability to lower blood sugar levels through a variety of pathways, including reducing the synthesis of glucose in the liver and enhancing peripheral glucose absorption.
The results presented here were in accordance with previous investigation which disclosed improved insulin sensitivity and enhanced glucose metabolism in diet-induced obese rats subjected to L-arginine treatment7. The modulation of glucometabolic enzymes by L-arginine offers mechanistic insights into its potential efficacy in addressing hyperglycaemia associated with diabetes. These findings propose that the supplementation of L-arginine stimulates glycolysis and augments glucose utilization in type 2 diabetes conditions. The L-arginine supplementation was shown to reduce adiposity and improve insulin sensitivity in animal models of obesity as well as in patients with diabetes and obesity47. Furthermore, investigations in rats have demonstrated that L-arginine can reduce plasma glucose levels, improving glucose tolerance48. On the other hand, metformin’s enduring benefits stem partly from its ability to counteract glycation, a process impairing protein function and fostering extracellular matrix buildup. Its composition facilitates direct binding to potent glycating agents like methylglyoxal49,50. This counters non-enzymatic glycation, pivotal in type 2 diabetes vascular complications. The concept of “metabolic memory” underscores metformin’s crucial role in mitigating long-term type 2 diabetes vascular risks51. Its effects extend beyond hyperglycemia, encompassing insulin resistance and ischemia. Metformin curtails advanced glycation end products (AGEs) both in vitro52 and in type 2 diabetes animals’ organs53.
Diabetes is closely linked to oxidative stress, where elevated reactive oxygen species (ROS) production contributes to the pathogenesis of the disease42. First off, oxidative stress markers were significantly lower in the group receiving metformin and L-arginine concurrently than in the group receiving therapy alone or in the control group. This implies that these two substances may have a synergistic anti-oxidative impact when used together. This effect may be related to the complementary ways in which these two chemicals scavenge free radicals and strengthen antioxidant defense systems. The intake of L-arginine has been shown to improve oxidative metabolism, through an enhanced mitochondrial function, eventually improving physical performance48.
The observed decline in MDA levels implies that L-arginine supplementation may alleviate lipid peroxidation, consequently diminishing oxidative damage. This finding aligned with Gambardella et al.54 research, reporting reduced oxidative stress markers in a mouse model of type 2 diabetes treated with L-arginine. The augmentation of SOD activity further underscores the antioxidant potential of L-arginine, as heightened SOD activity can fortify cellular defense against oxidative stress55. Moreover, metformin bolsters antioxidant defenses and activates AMP-kinase, pivotal in glucose and lipid metabolism26. Its efficacy against insulin resistance and hyperglycemia arises from its multifaceted actions23.
This study placed particular emphasis on investigating how L-arginine and metformin supplementation impact the histopathological examination of the kidneys in male Wistar rats with alloxan-induced diabetes. Given the propensity of diabetes to induce renal complications, exploring potential interventions like L-arginine and metformin supplementation becomes pivotal for comprehending therapeutic avenues. The histopathological analysis yielded valuable insights into the synergistic renal effects of L-arginine and metformin in the type 2 diabetes context. The observed enhancements in kidney histology after L-arginine and metformin supplementation indicate a potential protective role against the adverse effects of diabetes on renal tissues56. The alleviation of histopathological changes in the kidneys can be ascribed to the anti-inflammatory properties of both metformin and L-arginine. In diabetes, chronic inflammation significantly contributes to renal damage36 and the reduction in pro-inflammatory cytokines (TNF-α, IL-6, C-reactive protein and caspase 3) noted in this study may contribute to the preservation of renal tissue integrity.
The discovery of L-Arginine and metformin’s synergistic effects on oxidative stress, inflammation and glucometabolic enzymes in diabetic rats has interesting implications for diabetes management. This combo medication has the potential to improve diabetes management by targeting oxidative stress and inflammation, both of which contribute significantly to diabetic complications. The use of this synergy implies the possibility of establishing more effective therapy procedures that enhance glucose metabolism while lowering the risk of diabetes-related organ damage. Future studies should include clinical studies to confirm these findings in humans and discover the best dosing regimens. Furthermore, studying the molecular processes implicated in the synergistic action may provide new treatment options for diabetes and associated metabolic illnesses.
CONCLUSION
The current research clarifies the complex interactions between L-arginine and metformin supplementation in reducing oxidative stress, inflammation and dysregulation of the glucometabolic system in male Wistar rats with alloxan-induced diabetes. It is discovered to the best of our knowledge that the synergistic benefits of these supplements through thorough investigation, suggest their potential as adjuvant medicines in the management of problems associated with diabetes. The combination of these drugs holds great therapeutic promise, as seen by the reported regulation of important glucometabolic enzymes. The results highlight the significance of investigating alternate approaches to address the complex pathophysiology of diabetes.
SIGNIFICANCE STATEMENT
This study looks at the possible synergistic effects of L-arginine and metformin supplementation on oxidative stress, inflammation and glucometabolic enzyme activity in male Wistar rats with alloxan-induced diabetes. The combination of these two medicines, which have separate mechanisms for enhancing glucose metabolism and lowering oxidative damage, could provide a novel therapeutic approach to alleviating diabetes consequences. This study seeks to provide new insights into diabetes management techniques by understanding the interactions between L-arginine and metformin, potentially leading to improved clinical outcomes.
REFERENCES
- Ling, C. and T. Rönn, 2019. Epigenetics in human obesity and type 2 diabetes. Cell Metab., 29: 1028-1044.
- Inzucchi, S.E., 2013. Diagnosis of diabetes. N. Engl. J. Med., 368: 192-193.
- Witte, M.B., F.J. Thorton, U. Tantry and A. Barbul, 2002. L-arginine supplementation enhances diabetic wound healing: Involvement of the nitric oxide synthase and arginase pathways. Metabolism, 51: 1269-1273.
- Piatti, P.M., L.D. Monti, G. Valsecchi, F. Magni and E. Setola et al., 2001. Long-term oral L-arginine administration improves peripheral and hepatic insulin sensitivity in type 2 diabetic patients. Diabetes Care, 24: 875-880.
- Szkudelski, T., 2001. The mechanism of alloxan and streptozotocin action in B cells of the rat pancreas. Physiol. Res., 50: 537-546.
- Wu, G. and C.J. Meininger, 2008. Analysis of citrulline, arginine, and methylarginines using high-performance liquid chromatography. Methods Enzymol., 440: 177-189.
- Jobgen, W., C.J. Meininger, S.C. Jobgen, P. Li and M.J. Lee et al., 2009. Dietary L-arginine supplementation reduces white fat gain and enhances skeletal muscle and brown fat masses in diet-induced obese rats. J. Nutr., 139: 230-237.
- Lundberg, J.O. and E. Weitzberg, 2022. Nitric oxide signaling in health and disease. Cell, 185: 2853-2878.
- Adela, R., S.K. Nethi, P.K. Bagul, A.K. Barui and S. Mattapally et al., 2015. Hyperglycaemia enhances nitric oxide production in diabetes: A study from South Indian patients. PLoS ONE, 10.
- Narne, P., K.C. Ponnaluri, S. Singh, M. Siraj and M. Ishaq, 2013. Association of the genetic variants of endothelial nitric oxide synthase gene with angiographically defined coronary artery disease and myocardial infarction in South Indian patients with type 2 diabetes mellitus. J. Diabetes Complications, 27: 255-261.
- Ozcelik, O. and S. Algul, 2017. Nitric oxide levels in response to the patients with different stage of diabetes. Cell. Mol. Biol., 63: 49-52.
- Habib, S.S., K.A. Al-Regaiey, T. Al-Khlaiwi, S.M. Habib, S. Bashir, F. Al-Hussain and S.H. Habib, 2022. Serum inducible and endothelial nitric oxide synthase in coronary artery disease patients with type 2 diabetes mellitus. Eur. Rev. Med. Pharmacol. Sci., 26: 3695-3702.
- Bahadoran, Z., P. Mirmiran, K. Kashfi and A. Ghasemi, 2023. Vascular nitric oxide resistance in type 2 diabetes. Cell Death Dis., 14.
- Paulo, M., D.E.F.R. Costa, D. Bonaventura, C.N. Lunardi and L.M. Bendhack, 2020. Nitric oxide donors as potential drugs for the treatment of vascular diseases due to endothelium dysfunction. Curr. Pharm. Des., 26: 3748-3759.
- Bender, S.B., E.K. Herrick, N.D. Lott and R.E. Klabunde, 2007. Diet-induced obesity and diabetes reduce coronary responses to nitric oxide due to reduced bioavailability in isolated mouse hearts. Diabetes Obes. Metab., 9: 688-696.
- Suessenbacher, A., A. Lass, B. Mayer and F. Brunner, 2002. Antioxidative and myocardial protective effects of L-arginine in oxygen radical-induced injury of isolated perfused rat hearts. Naunyn-Schmiedeberg's Arch. Pharmacol., 365: 269-276.
- Forzano, I., R. Avvisato, F. Varzideh, S.S. Jankauskas and A. Cioppa et al., 2023. L-arginine in diabetes: Clinical and preclinical evidence. Cardiovasc. Diabetology, 22.
- Fu, W.J., T.E. Haynes, R. Kohli, J. Hu and W. Shi et al., 2005. Dietary L-arginine supplementation reduces fat mass in Zucker diabetic fatty rats. J. Nutr., 135: 714-721.
- Mendez, J.D. and F. Balderas, 2001. Regulation of hyperglycemia and dyslipidemia by exogenous L-arginine in diabetic rats. Biochimie, 83: 453-458.
- Mohan, I. and U. Cas, 1998. Effects of L-arginine-nitric oxide system on chemical induced diabetes mellitus. Free Radical Biol. Med., 25: 757-765.
- Knowler, W.C., E. Barrett-Connor, S.E. Fowler, R.F. Hamman and J.M. Lachin et al., 2002. Reduction in the incidence of type 2 diabetes with lifestyle intervention or metformin. N. Engl. J. Med., 346: 393-403.
- Yoshida, T., A. Okuno, J. Tanaka, K. Takahashi and R. Nakashima et al., 2009. Metformin primarily decreases plasma glucose not by gluconeogenesis suppression but by activating glucose utilization in a non-obese type 2 diabetes Goto-Kakizaki rats. Eur. J. Pharmacol., 623: 141-147.
- Kirpichnikov, D., S.I. McFarlane and J.R. Sowers, 2002. Metformin: An update. Ann. Intern. Med., 137: 25-33.
- Verma, S., S. Bhanot and J.H. McNeill, 1994. Antihypertensive effects of metformin in fructose-fed hyperinsulinemic, hypertensive rats. J. Pharmacol. Exp. Ther., 271: 1334-1337.
- Zou, M.H., S.S. Kirkpatrick, B.J. Davis, J.S. Nelson and W.G. Wiles et al., 2004. Activation of the AMP-activated protein kinase by the anti-diabetic drug metformin in vivo. J. Biol. Chem., 279: 43940-43951.
- Leverve, X.M., B. Guigas, D. Detaille, C. Batandier and E.A. Koceir et al., 2003. Mitochondrial metabolism and type-2 diabetes: A specific target of metformin. Diabetes Metab., 29: 6S88-6S94.
- UKPDS Group, 1998. Effect of intensive blood-glucose control with metformin on complications in overweight patients with type 2 diabetes (UKPDS 34). Lancet, 352: 854-865.
- Sartoretto, J.L., G.A.N. Melo, M.H.C. Carvalho, D. Nigro and R.T. Passaglia et al., 2005. Metformin treatment restores the altered microvascular reactivity in neonatal streptozotocin-induced diabetic rats increasing NOS activity, but not NOS expression. Life Sci., 77: 2676-2689.
- Romualdi, D., B. Costantini, L. Selvaggi, M. Giuliani and F. Cristello et al., 2008. Metformin improves endothelial function in normoinsulinemic PCOS patients: A new prospective. Hum. Reprod., 23: 2127-2133.
- Ighodaro, O.M., A.M. Adeosun and O.A. Akinloye, 2017. Alloxan-induced diabetes, a common model for evaluating the glycemic-control potential of therapeutic compounds and plants extracts in experimental studies. Medicina, 53: 365-374.
- Saka, W.A., T.E. Ayoade, T.M. Akhigbe and R.E. Akhigbe, 2021. Moringa oleifera seed oil partially abrogates 2,3-dichlorovinyl dimethyl phosphate (Dichlorvos)-induced cardiac injury in rats: Evidence for the role of oxidative stress. J. Basic Clin. Physiol. Pharmacol., 32: 237-246.
- Nwankwo, R.C., M.D. Ibegbu, K.C. Onyekwelu, C.S. Ejezie, J.E. Ikekpeazu and F.E. Ejezie, 2019. Biochemical and histopathological effects of sub-acute exposure of albino rats to fumigants-dichlorvos and cypermethrin. Interdiscip. Toxicol., 12: 180-185.
- Igbayilola, Y.D. and M.G. Gujja, 2024. Alpha-amylase and alpha-glucosidase upregulated glucose homeostasis in high-fat fed Wistar rats supplemented with cocoa flavonoid-rich aqueous. Food Biosci., 59.
- Dimeji, I.Y., A.O. Samson, A. Kayode, A. Ikponwosa, W.O. Dada and O.F. Adesina, 2023. Glucometabolic response to walnut (Juglans regia L.) supplementation during gestation and/or lactation in offspring of Sprague-Dawley rats. Singapore J. Sci. Res., 13: 79-87.
- Vilela, D.D., L.G. Peixoto, R.R. Teixeira, N.B. Baptista and D.C. Caixeta et al., 2016. The role of metformin in controlling oxidative stress in muscle of diabetic rats. Oxid. Med. Cell. Longevity, 2016.
- Aebi, H., 1984. Catalase in vitro. Methods Enzymol., 105: 121-126.
- Uchiyama, M. and M. Mihara, 1978. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem., 86: 271-278.
- Shaheen, M.Y., A.M. Basudan, A.A. Niazy, J.J.J.P. van den Beucken, J.A. Jansen and H.S. Alghamdi, 2021. Histological and histomorphometric analyses of bone regeneration in osteoporotic rats using a xenograft material. Materials, 14.
- Donath, M.Y. and S.E. Shoelson, 2011. Type 2 diabetes as an inflammatory disease. Nat. Rev. Immunol., 11: 98-107.
- Karbach, S., P. Wenzel, A. Waisman, T. Munzel and A. Daiber, 2014. eNOS uncoupling in cardiovascular diseases-the role of oxidative stress and inflammation. Curr. Pharm. Des., 20: 3579-3594.
- Morris Jr., S.M., 2016. Arginine metabolism revisited. J. Nutr., 146: 2579S-2586S.
- Deanfield, J.E., J.P. Halcox and T.J. Rabelink, 2017. Endothelial function and dysfunction: Testing and clinical relevance. Circulation, 115: 1285-1295.
- Kim, F., M. Pham, E. Maloney, N.O. Rizzo and G.J. Morton et al., 2008. Vascular inflammation, insulin resistance, and reduced nitric oxide production precede the onset of peripheral insulin resistance. Arterioscler., Thromb., Vasc. Biol., 28: 1982-1988.
- Morin-Papunen, L., K. Rautio, A. Ruokonen, P. Hedberg, M. Puukka and J.S. Tapanainen, 2003. Metformin reduces serum C-reactive protein levels in women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab., 88: 4649-4654.
- Caballero, A.E., A. Delgado, C.A. Aguilar-Salinas, A.N. Herrera and J.L. Castillo et al., 2004. The differential effects of metformin on markers of endothelial activation and inflammation in subjects with impaired glucose tolerance: A placebo-controlled, randomized clinical trial. J. Clin. Endocrinol. Metab., 89: 3943-3948.
- Isoda, K., J.L. Young, A. Zirlik, L.A. MacFarlane and N. Tsuboi et al., 2006. Metformin inhibits proinflammatory responses and nuclear factor-κB in human vascular wall cells. Arterioscler., Thromb., Vasc. Biol., 26: 611-617.
- McKnight, J.R., F.C. Satterfield, W.S. Jobgen, S.B. Smith and T.E. Spencer et al., 2010. Beneficial effect of L-arginine on reducing obesity: Potential mechanisms and important implications for human health. Amino Acids, 39: 349-357.
- Newsholme, P., K.N. Keane, R. Carlessi and V. Cruzat, 2019. Oxidative stress pathways in pancreatic β-cells and insulin-sensitive cells and tissues: Importance to cell metabolism, function, and dysfunction. Am. J. Physiol. Cell Physiol., 317: C420-C433.
- Beisswenger, P. and D. Ruggiero-Lopez, 2003. Metformin inhibition of glycation processes. Diabetic Metab., 29: 6S95-103.
- Ihnat, M.A., J.E. Thorpe and A. Ceriello, 2007. Hypothesis: The ‘metabolic memory’, the new challenge of diabetes. Diabetic Med., 24: 582-586.
- Kiho, T., M. Kato, S. Usui and K. Hirano, 2005. Effect of buformin and metformin on formation of advanced glycation end products by methylglyoxal. Clin. Chim. Acta, 358: 139-145.
- Ruggiero-Lopez, D., M. Lecomte, G. Moinet, G. Patereau, M. Lagarde and N. Wiernsperger, 1999. Reaction of metformin with dicarbonyl compounds. Possible implication in the inhibition of advanced glycation end product formation. Biochem. Pharmacol., 58: 1765-1773.
- Tanaka, Y., H. Uchino, T. Shimizu, H. Yoshii and M. Niwa et al., 1999. Effect of metformin on advanced glycation endproduct formation and peripheral nerve function in streptozotocin-induced diabetic rats. Eur. J. Pharmacol., 376: 17-22.
- Gambardella, J., A. Fiordelisi, L. Spigno, L. Boldrini and G. Lungonelli et al., 2021. Effects of chronic supplementation of L-arginine on physical fitness in water polo players. Oxid. Med. Cell. Longevity.
- Dubey, H., A. Dubey, K. Gulati and A. Ray, 2022. Protective effects of L-arginine on cognitive deficits and biochemical parameters in an experimental model of type-2 diabetes mellitus induced Alzheimer's disease in rats. J. Physiol. Pharmacol., 73: 3-17.
- Reutens, A.T. and R.C. Atkins, 2011. Epidemiology of Diabetic Nephropathy. In: Contributions to Nephrology, Lai, K.N. and S.C.W. Tang (Eds.), Karger, Switzerland, ISBN: 978-3-8055-9743-2, pp: 1-7.
How to Cite this paper?
APA-7 Style
Adeoye,
S.W., Dimeji,
I.Y., Jabba,
L.H., Emmanuel,
O., Samuel,
A.T., Oluwakemi,
O.D., Muhammed,
Z. (2024). Synergistic Effects of L-Arginine and Metformin on Oxidative Stress, Inflammation and Glucometabolic Enzymes in Diabetic Rats. Trends in Applied Sciences Research, 19(1), 180-198. https://doi.org/10.3923/tasr.2024.180.198
ACS Style
Adeoye,
S.W.; Dimeji,
I.Y.; Jabba,
L.H.; Emmanuel,
O.; Samuel,
A.T.; Oluwakemi,
O.D.; Muhammed,
Z. Synergistic Effects of L-Arginine and Metformin on Oxidative Stress, Inflammation and Glucometabolic Enzymes in Diabetic Rats. Trends Appl. Sci. Res 2024, 19, 180-198. https://doi.org/10.3923/tasr.2024.180.198
AMA Style
Adeoye
SW, Dimeji
IY, Jabba
LH, Emmanuel
O, Samuel
AT, Oluwakemi
OD, Muhammed
Z. Synergistic Effects of L-Arginine and Metformin on Oxidative Stress, Inflammation and Glucometabolic Enzymes in Diabetic Rats. Trends in Applied Sciences Research. 2024; 19(1): 180-198. https://doi.org/10.3923/tasr.2024.180.198
Chicago/Turabian Style
Adeoye, Saka, Waidi, Igbayilola Yusuff Dimeji, Lawan, Hamidu Jabba, Osukoya Emmanuel, Abaloja Temiloluwa Samuel, Ogunsina Damola Oluwakemi, and Zakari, Muhammed.
2024. "Synergistic Effects of L-Arginine and Metformin on Oxidative Stress, Inflammation and Glucometabolic Enzymes in Diabetic Rats" Trends in Applied Sciences Research 19, no. 1: 180-198. https://doi.org/10.3923/tasr.2024.180.198

This work is licensed under a Creative Commons Attribution 4.0 International License.